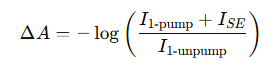ในบทความก่อนหน้านี้ เราได้ตรวจสอบกระบวนการพื้นฐานและหลักการโดยละเอียดของการตรวจจับสเปกโทรสโกปีการดูดกลืนแสงชั่วคราวอย่างละเอียด ในบทความนี้ เราจะสำรวจเพิ่มเติมเกี่ยวกับการประยุกต์ใช้สเปกโทรสโกปีการดูดกลืนแสงชั่วคราวที่เร็วเป็นพิเศษในระบบโมเลกุล โดยมุ่งเน้นไปที่ทั้งการดูดกลืนแสงชั่วคราวที่เร็วเป็นพิเศษและสเปกโทรสโกปีการสะท้อนกลับชั่วคราวที่เร็วเป็นพิเศษ
01 สภาวะตื่นเต้นระดับโมเลกุล (การฟอกสีแบบพื้นดิน)
เรามาเริ่มด้วยตัวอย่างง่ายๆ โดยใช้โมเลกุลอินทรีย์ C และกระบวนการสถานะตื่นเต้นขั้นพื้นฐานที่สุด S₁ เพื่อแสดงหลักการเบื้องหลังการสร้างสเปกตรัมการดูดกลืนแสงชั่วคราว
รูปที่ 1a แสดงสเปกตรัมการดูดกลืนแสงในสถานะคงตัวของโมเลกุล C ในสถานะพื้น (S₀) เมื่อโมเลกุล C ถูกตื่นเต้น (ถูกสูบ) ด้วยเลเซอร์แบบพัลซิ่ง (สเปกโทรสโกปีการดูดกลืนแสงชั่วคราวต้องใช้แหล่งกำเนิดแสงแบบพัลส์เพื่อการกระตุ้น) ส่วนหนึ่งของโมเลกุลจะดูดซับโฟตอนและผ่านการเปลี่ยนแปลงทางอิเล็กทรอนิกส์ S₀ → S₁ ทำให้เกิดโมเลกุลสถานะตื่นเต้น C* (รูปที่ 1b) ณ จุดนี้ ตัวอย่างประกอบด้วยส่วนผสมของโมเลกุลสถานะพื้นดินที่ไม่ถูกกระตุ้น (C) และโมเลกุลที่มีสถานะตื่นเต้น (C*) สัดส่วนของ C* ขึ้นอยู่กับความยาวคลื่นของปั๊มและความเข้มของแหล่งกำเนิดแสงกระตุ้น
จากนั้น หากเราตรวจสอบสเปกตรัมการดูดกลืนแสงทันทีของตัวอย่างในเวลาหน่วงเฉพาะ (ก่อนที่ C* จะสลายตัวกลับสู่สถานะพื้น) เราจะสังเกตได้ว่าสเปกตรัมการดูดกลืนแสง A C+C* ของตัวอย่างที่ตื่นเต้น นั้นอ่อนกว่าความเข้มข้นเดียวกันของโมเลกุลสถานะพื้น A C (ดังแสดงในรูปที่ 1a) เนื่องจากโมเลกุลในสถานะตื่นเต้น C* ไม่แสดงคุณลักษณะการดูดกลืนแสงแบบเดียวกับโมเลกุลในสถานะพื้น C—เช่น AC ≠ A C* . การเปลี่ยนแปลงในการดูดซับก่อนและหลังการกระตุ้นนี้ก่อให้เกิดพื้นฐานของสเปกโทรสโกปีการดูดกลืนแสงชั่วคราว
ดังนั้นการดูดกลืนแสงที่ลดลงที่สังเกตได้ในรูปที่ 1a หลังจากการกระตุ้นนั้นเกิดจากการมีอยู่ของโมเลกุลที่มีสถานะตื่นเต้น C* ดังที่แสดงในรูปที่ 1c โดยการลบ AC ออก จาก A C+C* เราจะได้สเปกตรัมการดูดกลืนแสงชั่วคราว (∆A) ของโมเลกุล C ณ จุดเวลาที่กำหนดหลังการกระตุ้น โดยทั่วไปสเปกตรัมดิฟเฟอเรนเชียล ∆A จะแสดงเป็น: ∆A = A ปั๊ม – อัน ปั๊ม นั่นคือการเปลี่ยนแปลงในการดูดซับก่อนและหลังการกระตุ้น ตามหลักการนี้ สัญญาณ ∆A ที่เป็นลบซึ่งสอดคล้องกับจุดสูงสุดของการดูดกลืนแสงในสภาวะคงตัว บ่งชี้ถึงสารฟอกขาวในสถานะพื้นดิน (GSB)
โดยพื้นฐานแล้ว กระบวนการใดๆ ที่การกระตุ้นด้วยแสงทำให้โมเลกุลออกจากสถานะพื้นจะส่งผลให้เกิดสัญญาณการฟอกขาวในสถานะพื้น ด้วยการปรับการหน่วงเวลาระหว่างพัลส์ของปั๊มและโพรบโดยใช้เทคนิคที่เหมาะสม เราจะได้สเปกตรัมการดูดกลืนแสงชั่วคราวที่แก้ไขตามเวลา ∆A(t) ของโมเลกุล C (ดังแสดงในรูปที่ 1d) เนื่องจากการกระตุ้นเกิดขึ้นในช่วงเวลาต่ำกว่าเฟมโตวินาที สัญญาณ ∆A ที่สอดคล้องกันจะถูกสร้างขึ้นทันทีหลังจากการกระตุ้น อย่างไรก็ตาม ในทางปฏิบัติ การสร้างสัญญาณ ∆A ถูกจำกัดโดยฟังก์ชันการตอบสนองของเครื่องมือ (IRF)
ตามกระบวนการทางแสงที่แสดงในรูปที่ 1b ชุดสเปกตรัมนี้สะท้อนการสลายตัวของโมเลกุลที่มีสถานะตื่นเต้น C* กลับสู่สถานะพื้นผ่านวิถีการแผ่รังสี (K r ) หรือไม่เป็นรังสี (K nr ) ในทำนองเดียวกัน สัญญาณ GSB จะปรากฏขึ้นทันทีเมื่อมีการกระตุ้น และค่อยๆ ลดลงเมื่อเวลาผ่านไป
จากสเปกตรัม ∆A(t) เราสามารถแยกเส้นโค้งจลน์ศาสตร์ชั่วคราวที่ความยาวคลื่นจำเพาะได้ (รูปที่ 1d) ขอบที่เพิ่มขึ้นอย่างรวดเร็วสะท้อนถึงกระบวนการกระตุ้น ในขณะที่การฟื้นตัวเป็นศูนย์บ่งชี้ถึงการสลายตัวของสภาวะตื่นเต้น เมื่อปรับเส้นโค้งนี้ให้เหมาะสม เราจะได้ค่าพารามิเตอร์จลน์ศาสตร์ เช่น อัตราการสลายตัวของสถานะตื่นเต้น
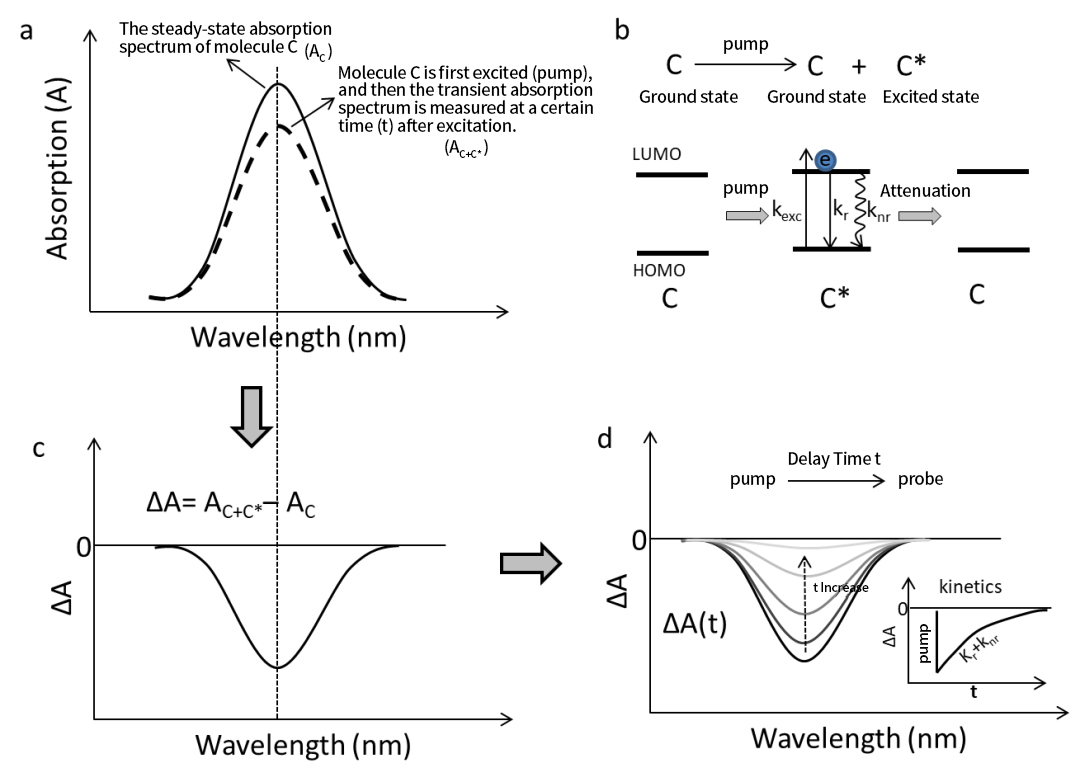
รูปที่ 1 กระบวนการสร้างสเปกตรัมการดูดกลืนแสงชั่วคราวโดยใช้โมเลกุล C เป็นตัวอย่าง
(a) การเปรียบเทียบระหว่างสเปกตรัมการดูดกลืนแสงในสภาวะคงตัวของโมเลกุลกับสเปกตรัมการดูดกลืนแสง ณ เวลาที่กำหนดหลังการกระตุ้น (b) กระบวนการเปลี่ยนผ่านทางอิเล็กทรอนิกส์ขั้นพื้นฐานหลังจากการกระตุ้นของโมเลกุล C หลังจากที่โมเลกุล C ถูกตื่นเต้น ส่วนหนึ่งของโมเลกุล C จะตื่นเต้นและเปลี่ยนเป็นโมเลกุลที่มีสถานะตื่นเต้น (C*) kr และ knr คืออัตราการสลายตัวของการแผ่รังสีและไม่มีการแผ่รังสีของโมเลกุลสถานะตื่นเต้น C* ตามลำดับ (c) สเปกตรัมการดูดกลืนแสงชั่วคราว ∆A ณ เวลาที่กำหนดหลังจากการกระตุ้นของโมเลกุล C ซึ่งได้มาจากการลบสเปกตรัมการดูดกลืนแสงสถานะพื้น (A C ) ออกจากสเปกตรัมการดูดกลืนแสง (AC + C* ) ในเวลาที่กำหนดหลังการกระตุ้น (d) โดยการรวบรวมสเปกตรัมการดูดกลืนแสงชั่วคราว (∆A) ในเวลาต่าง ๆ (t) หลังจากการกระตุ้นของโมเลกุล C กระบวนการวิวัฒนาการชั่วคราวของสถานะตื่นเต้นของโมเลกุล C และจลนศาสตร์การผ่อนคลายที่ความยาวคลื่นจำเพาะสามารถกำหนดได้
จากคำอธิบายข้างต้น เราได้เรียนรู้หลักการพื้นฐานเบื้องหลังการสร้างและการตรวจจับสเปกตรัมการดูดกลืนแสงชั่วคราวสำหรับโมเลกุล C อย่างไรก็ตาม ในการใช้งานจริง นอกเหนือจากสัญญาณฟอกขาวในสถานะพื้นดิน (GSB) ที่กล่าวถึงก่อนหน้านี้แล้ว สเปกตรัมการดูดกลืนแสงชั่วคราวยังมีสัญญาณลักษณะเฉพาะอื่นๆ ที่สะท้อนถึงไดนามิกของสถานะตื่นเต้น C*
ตัวอย่างเช่น โมเลกุลที่มีสถานะตื่นเต้น C* สามารถผ่านกระบวนการดูดซับโฟตอนแบบใหม่ ซึ่งเรียกว่าการดูดซับโดยสภาวะตื่นเต้น (ESA)
ดังที่แสดงในรูปที่ 2a สถานะ S₁ สามารถดูดซับพลังงานโฟตอนเพิ่มเติมเพื่อเปลี่ยนไปสู่สถานะตื่นเต้นที่สูงขึ้น (Sₙ) ดังนั้น หลังจากที่ตัวอย่างตื่นเต้น ไฟโพรบจึงสามารถตรวจจับสัญญาณการดูดซับในสถานะตื่นเต้นใหม่ได้ สัญญาณ ESA เหล่านี้ปรากฏเป็นสัญญาณเชิงบวกในสเปกตรัมการดูดกลืนแสงชั่วคราว (รูปที่ 2b) เนื่องจากระดับพลังงานของการเปลี่ยนสถานะตื่นเต้นมักจะกระจายไปในช่วงกว้าง สัญญาณ ESA ที่ได้จึงมักจะครอบคลุมช่วงความยาวคลื่นที่กว้าง (รูปที่ 2b)
สิ่งสำคัญคือต้องสังเกตว่าช่วงสเปกตรัมและความเข้มของ ESA อาจแตกต่างกันอย่างมากในโมเลกุลต่างๆ หาก ESA ซ้อนทับกับสัญญาณ GSB สเปกตรัมการดูดกลืนแสงชั่วคราวที่เกิดขึ้นอาจปรากฏขึ้นดังแสดงในรูปที่ 2ค กระบวนการสลายสถานะตื่นเต้นที่แสดงในรูปที่ 1b สอดคล้องกับวิวัฒนาการชั่วคราวของสเปกตรัมชั่วคราวที่แสดงในรูปที่ 2d ซึ่งทั้งสารฟอกขาวในสถานะพื้นดินและสัญญาณการดูดซับในสถานะตื่นเต้นลดลงเมื่อเวลาผ่านไป
เนื่องจากทั้งสัญญาณ GSB และ ESA มาจากสถานะตื่นเต้นเดียวกัน เราอาจสังเกตเห็นจุดไอออสเบสติกในสเปกตรัมชั่วคราวของรูปที่ 2d ซึ่งเป็นความยาวคลื่นที่การดูดกลืนแสงยังคงคงที่เมื่อเวลาผ่านไป สิ่งนี้บ่งชี้ว่าการเปลี่ยนแปลงในการดูดกลืนแสงที่ด้านใดด้านหนึ่งของจุดนี้เกิดจากกระบวนการทางแสงแบบเดียวกัน กล่าวคือ การเสื่อมสลายของสภาวะตื่นเต้น
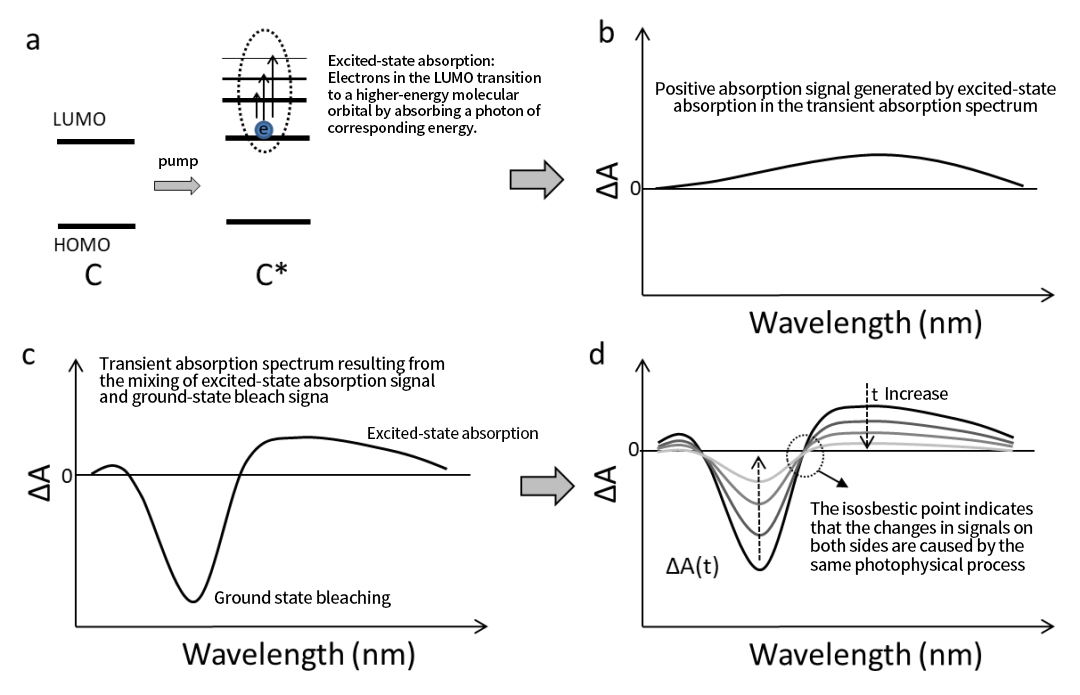
รูปที่ 2 (a) กระบวนการดูดซับสถานะตื่นเต้นของโมเลกุล C (b) สัญญาณสเปกโทรสโกปีการดูดกลืนแสงชั่วคราวที่เกิดจากการดูดกลืนสถานะตื่นเต้นของโมเลกุล C (c) สัญญาณสเปกโทรสโกปีการดูดกลืนแสงชั่วคราวหลังจากผสมสัญญาณการดูดกลืนแสงสถานะตื่นเต้นกับสัญญาณฟอกขาวสถานะพื้นดิน (d) วิวัฒนาการเวลาของสเปกตรัมการดูดกลืนแสงชั่วคราวที่รวมเอาการดูดซับในสถานะตื่นเต้นและการฟอกขาวในสถานะพื้นดิน
02 สถานะ Triplet โมเลกุล
Transient Absorption Spectroscopy ยังสามารถใช้เพื่อสังเกตกระบวนการข้ามระบบ (ISC) ระหว่างสถานะสายเดี่ยวของโมเลกุลและสถานะแฝด
รูปที่ 3a แสดงให้เห็นถึงกระบวนการ ISC โดยที่ K₀(S) และ K₀(T) เป็นตัวแทนของค่าคงที่อัตราการสลายตัวของสถานะตื่นเต้นของเสื้อกล้ามและสถานะแฝดตามลำดับ ซึ่งรวมถึงวิถีการสลายตัวทั้งหมด ทั้งแบบแผ่รังสีและแบบไม่แผ่รังสี ตราบใดที่โมเลกุลยังไม่กลับสู่สถานะพื้น สัญญาณสารฟอกขาวในสถานะพื้น (GSB) จะยังคงมีอยู่ ไม่ว่าโมเลกุลจะอยู่ในสถานะเสื้อกล้ามหรือแฝดก็ตาม ดังนั้นธนาคารออมสินเพียงอย่างเดียวจึงไม่สามารถให้ข้อมูลเกี่ยวกับการเปลี่ยนแปลงระหว่างสถานะที่ตื่นเต้นได้
อย่างไรก็ตาม เราสามารถดึงข้อมูลจลนศาสตร์ของสถานะทริปเล็ตผ่านการดูดซับสถานะตื่นเต้น (ESA) ของสถานะทริปเล็ต ซึ่งเป็นไปตามหลักการเดียวกันกับ ESA เสื้อกล้าม หากต้องการใช้สเปกโทรสโกปีการดูดกลืนแสงชั่วคราวเพื่อศึกษาสถานะทริปเล็ต ต้องเป็นไปตามเงื่อนไขสองประการ:
1. อัตรา ISC จากเสื้อกล้ามถึงแฝดจะต้องสูงเพียงพอ—เทียบได้กับหรือเร็วกว่าอัตราการสลายตัวของสถานะเสื้อกล้าม K₀(S)—มิฉะนั้น สถานะแฝดจะไม่ก่อตัว
2. สัญญาณ ESA ของสถานะแฝดจะต้องอยู่ภายในช่วงสเปกตรัมที่ตรวจพบได้
รูปที่ 3b แสดงสเปกตรัมการดูดกลืนแสงชั่วคราวทั่วไปสำหรับกระบวนการ ISC สายเดี่ยวถึงสาม สัญญาณ ESA ของสถานะแฝดซ้อนทับกับสัญญาณ GSB บางส่วน และสันนิษฐานว่า K ISC ≫ K₀(S)
เมื่อเวลาหน่วงของปั๊ม-โพรบ (t) เพิ่มขึ้น เราจะสังเกตสัญญาณ ESA แฝดที่ก่อตัวและค่อยๆ เข้มข้นขึ้น ซึ่งสะท้อนถึงจลนศาสตร์ของกระบวนการ ISC ถ้า K ISC ≫ K₀(S) ดังนั้นสัญญาณ GSB ควรจะคงไว้ไม่เปลี่ยนแปลงจนกว่าสถานะแฝดจะสลายกลับไปเป็นสถานะพื้น อย่างไรก็ตาม ในรูปที่ 3b เราสังเกตเห็นการสลายตัวเล็กน้อยและการเปลี่ยนแปลงสเปกตรัมในสัญญาณ GSB นี่ไม่ได้เกิดจากการเปลี่ยนแปลงที่เกิดขึ้นจริงในสารฟอกขาวในสถานะพื้น แต่เกิดจากการเติบโตของสัญญาณ ESA บวกที่ทับซ้อนกันจากสถานะแฝด
การรบกวนสัญญาณประเภทนี้พบได้ทั่วไปในการทดลองการดูดกลืนแสงชั่วคราว และต้องพิจารณาอย่างรอบคอบในระหว่างการวิเคราะห์ข้อมูล เทคนิคต่างๆ เช่น การปรับให้เหมาะสมโดยรวมหรือการสลายตัวของค่าเอกพจน์ (SVD) มักใช้เพื่อแยกและตีความสัญญาณที่ทับซ้อนกัน
เมื่อเวลาหน่วง t ยังคงเพิ่มขึ้น ทั้งสัญญาณ ESA แฝดและสัญญาณ GSB สลายตัวจนเหลือศูนย์ในที่สุด ซึ่งสะท้อนถึงจลนศาสตร์การสลายตัวของสถานะแฝด นั่นคือ K₀(T) เนื่องจากธรรมชาติของการสลายตัวของแฝดที่ห้ามการหมุน กระบวนการนี้จึงมักเกิดขึ้นในช่วงเวลาที่ค่อนข้างยาว
จากสเปกตรัม ∆A(t) สามารถดึงเส้นโค้งจลนศาสตร์ชั่วคราวที่ความยาวคลื่นเฉพาะภายในขอบเขต ESA แฝด (ดูรูปที่ 3b) และพารามิเตอร์จลน์ศาสตร์เช่น K ISC และ K₀(T) สามารถรับได้ผ่านการปรับเส้นโค้ง
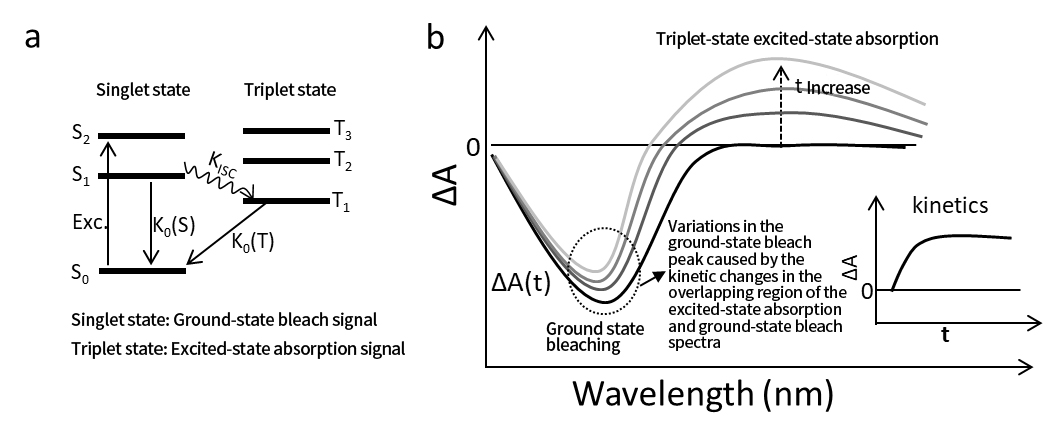
รูปที่ 3 (a) กระบวนการไดนามิก เช่น การข้ามระบบระหว่างกัน (ISC) และการสลายตัวระหว่างสถานะเสื้อกล้ามและแฝดของโมเลกุล ในสเปกโทรสโกปีการดูดกลืนแสงชั่วคราว พลวัตของสถานะสายเดี่ยวและสถานะแฝดสามารถจับได้ผ่านการฟอกขาวในสถานะพื้นและสัญญาณการดูดกลืนแสงในสถานะแฝดที่ตื่นเต้น (b) วิวัฒนาการชั่วคราวของสเปกตรัมการดูดกลืนแสงชั่วคราว ซึ่งแสดงให้เห็นกระบวนการ ISC และเส้นโค้งจลน์ของสถานะแฝด
03 การถ่ายโอนอิเล็กตรอนด้วยแสง
การถ่ายโอนอิเล็กตรอนด้วยแสงเป็นกระบวนการจลนศาสตร์ที่สำคัญอย่างยิ่งในระบบการแปลงด้วยแสง และแสดงถึงกลไกหลักในอุปกรณ์ต่างๆ เช่น เซลล์แสงอาทิตย์ ตัวเร่งปฏิกิริยาด้วยแสง และเครื่องตรวจจับแสง
สเปกโทรสโกปีการดูดกลืนแสงชั่วคราวถือได้ว่าเป็นหนึ่งในเทคนิคที่มีประสิทธิผลมากที่สุดในการตรวจสอบกระบวนการถ่ายโอนอิเล็กตรอนที่เกิดจากแสง ไม่ว่าจะเกิดขึ้นระหว่างวัสดุที่แตกต่างกันหรือภายในวัสดุเดียว
ในบทความนี้ เราเริ่มต้นด้วยการแสดงให้เห็นว่าสามารถใช้สเปกโทรสโกปีการดูดกลืนแสงชั่วคราวเพื่อศึกษากระบวนการถ่ายโอนอิเล็กตรอนระหว่างโมเลกุลชนิดต่างๆ ได้อย่างไร กระบวนการถ่ายโอนอิเล็กตรอนหรือประจุในระบบอื่น เช่น ระบบเซมิคอนดักเตอร์หรือระบบไฮบริดเซมิคอนดักเตอร์/โมเลกุล จะถูกกล่าวถึงในหัวข้อต่อๆ ไป
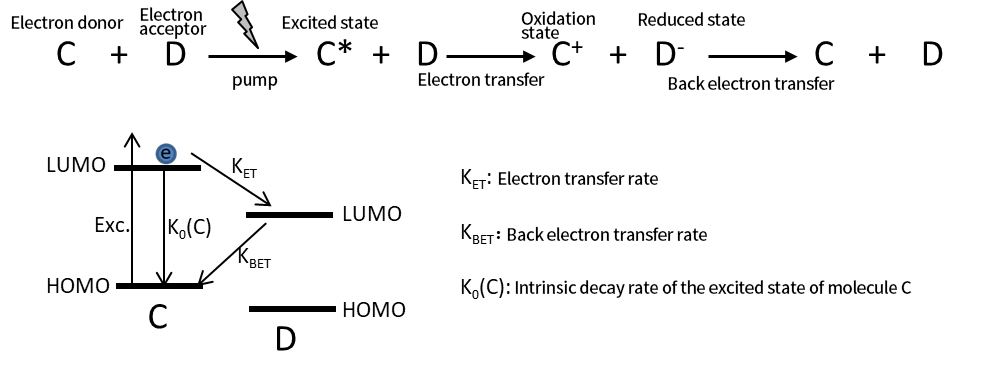
รูปที่ 4 กระบวนการปฏิกิริยาการถ่ายโอนอิเล็กตรอนด้วยแสงระหว่างโมเลกุล C และ D พร้อมกับการเปลี่ยนผ่านทางอิเล็กทรอนิกส์ระหว่างวงโคจรของโมเลกุลที่สอดคล้องกัน K0(C) แสดงถึงผลรวมของวิถีการสลายตัวแบบแผ่รังสีและแบบไม่แผ่รังสี
รูปที่ 4 แสดงกระบวนการถ่ายโอนอิเล็กตรอนด้วยแสงระหว่างโมเลกุลสองโมเลกุล ได้แก่ C และ D โมเลกุล C ทำหน้าที่เป็นผู้บริจาคอิเล็กตรอน และโมเลกุล D เป็นตัวรับอิเล็กตรอน เมื่อกระตุ้นด้วยแสง โมเลกุล C จะเข้าสู่สภาวะตื่นเต้นและถ่ายโอนอิเล็กตรอนไปยังโมเลกุล D หลังจากการถ่ายโอน C และ D จะกลายเป็นอนุมูลที่ถูกออกซิไดซ์ C⁺ และอนุมูล D⁻ แบบรีดิวซ์ ตามลำดับ
หากไม่มีปฏิกิริยาใดเกิดขึ้นอีก อิเล็กตรอนที่ถูกถ่ายโอนจะกลับสู่โมเลกุล C ในที่สุดโดยการถ่ายโอนอิเล็กตรอนด้านหลัง (BET) เพื่อฟื้นฟูระบบกลับสู่สถานะเริ่มต้น
โดยทั่วไปการถ่ายโอนอิเล็กตรอนด้านหลังจะเกิดขึ้นช้ากว่าการถ่ายโอนอิเล็กตรอนไปข้างหน้ามาก ซึ่งเป็นที่ต้องการในโฟโตคะตะไลซิสและเซลล์แสงอาทิตย์ การเดิมพันที่ช้ากว่าจะทำให้ C⁺ และ D⁻ อยู่ได้นานขึ้น ทำให้พวกมันมีส่วนร่วมในปฏิกิริยาเร่งปฏิกิริยาอื่น ๆ (เช่นในโฟโตคะตะไลซิส) หรืออำนวยความสะดวกในการสกัดและส่งออกประจุ (เช่นในเซลล์แสงอาทิตย์)
ในสเปกโทรสโกปีการดูดกลืนแสงชั่วคราว สิ่งสำคัญคือต้องเข้าใจว่าจากจุดยืนของโมเลกุล C⁺ และ D⁻ เป็นสปีชีส์ที่แตกต่างกันโดยมีลักษณะการดูดกลืนแสงแตกต่างจาก C และ D ดังนั้น ในระบบโมเลกุล สเปกตรัมการดูดกลืนแสงของผู้ให้และตัวรับจะเปลี่ยนไปหลังจากการถ่ายโอนอิเล็กตรอน ไม่เหมือนในเซมิคอนดักเตอร์ (เช่น จุดควอนตัม) โดยที่สเปกตรัมก่อนและหลังการถ่ายโอนประจุอาจไม่แตกต่างกันอย่างมีนัยสำคัญ สเปกตรัมการดูดกลืนแสงของอนุมูล C⁺ และ D⁻ สามารถกำหนดได้โดยวิธีเคมีไฟฟ้าร่วมกับการดูดกลืนในสภาวะคงตัว หรือผ่านสเปกโทรสโกปีการดูดกลืนแสงชั่วคราว ในระบบที่เกิดการถ่ายโอนประจุ
เราอธิบายสองสถานการณ์เพื่อแสดงให้เห็นว่าสเปกโทรสโกปีการดูดกลืนแสงชั่วคราวตรวจจับการถ่ายโอนอิเล็กตรอนที่เกิดจากแสงได้อย่างไร:
1) สเปกตรัมการดูดกลืนแสงสถานะคงตัวของ C และ D เป็นที่รู้จักและอยู่ภายในช่วงการตรวจจับ แต่สเปกตรัม C⁺ และ D⁻ ไม่เป็นที่รู้จักหรืออยู่นอกช่วงการตรวจจับ
รูปที่ 5a แสดงสเปกตรัมการดูดกลืนแสงในสภาวะคงตัวของ C และ D และสเปกตรัมการดูดกลืนแสงของตัวอย่างในเวลาที่กำหนดหลังจากที่ C รู้สึกตื่นเต้นและผ่านการถ่ายโอนอิเล็กตรอน หลังจากการถ่ายโอนอิเล็กตรอน ความเข้มของการดูดซับของ C และ D จะลดลง—คล้ายกับสถานการณ์ในรูปที่ 1a—เนื่องจากส่วนหนึ่งของโมเลกุลกลายเป็น C⁺ และ D⁻ ซึ่งมีลักษณะการดูดซับที่แตกต่างกัน
หากอัตราการถ่ายโอนอิเล็กตรอน (K ET ) เร็วกว่าอัตราการสลายตัวในสภาวะตื่นเต้นภายในของ C (K₀(C)) มาก ดังนั้น C* จะสลายตัวโดยการถ่ายโอนอิเล็กตรอนเป็นหลัก ในกรณีนี้ วิวัฒนาการชั่วคราวของสเปกตรัมการดูดกลืนแสงชั่วคราวจะปรากฏดังในรูปที่ 5b เมื่อเวลาหน่วงของปั๊มและโพรบเพิ่มขึ้น สัญญาณสารฟอกขาวสถานะพื้นดิน (GSB) ของ D จะค่อยๆ ปรากฏขึ้น (ตามรูปแบบ D⁻ ซึ่งลดจำนวนประชากร D) สะท้อนถึงจลนพลศาสตร์การถ่ายโอนอิเล็กตรอนจาก C ถึง D
ในเวลาเดียวกัน GSB ของ C จะปรากฏขึ้นทันทีหลังจากการกระตุ้นแต่ยังคงไม่เปลี่ยนแปลงเมื่อเวลาผ่านไป (ภายใต้ K ET ≫ K₀(C)) เนื่องจาก C* เปลี่ยนเป็น C⁺ โดยไม่กลับสู่สถานะพื้น หากมองเห็นการดูดกลืนสภาวะตื่นเต้น (ESA) ของ C เช่นกัน สัญญาณนี้จะปรากฏขึ้นครั้งแรกหลังจากการกระตุ้น จากนั้นสลายตัวเมื่อ C* เปลี่ยนเป็น C⁺ ดังนั้นการสลายตัวของสัญญาณ ESA จึงสามารถใช้เพื่อติดตามกระบวนการถ่ายโอนอิเล็กตรอนได้
หมายเหตุ: หาก K ET เทียบได้กับ K₀(C) GSB ของ C จะสลายตัวก่อนที่ BET จะเกิดขึ้น และ ESA ของ C จะสะท้อนทั้งการถ่ายโอนอิเล็กตรอนและการสลายตัวภายใน
เมื่อเวลาหน่วงเพิ่มขึ้นอีก กระบวนการถ่ายโอนอิเล็กตรอนด้านหลังจะเริ่มมีอิทธิพลเหนือ สัญญาณ GSB ของทั้ง C และ D เริ่มสลายตัวและหายไปในที่สุด ทำให้ระบบกลับสู่สถานะเริ่มต้น (รูปที่ 5c)
รูปที่ 5d แสดงเส้นโค้งจลน์ที่ดึงมาจากคุณลักษณะสเปกตรัมต่างๆ ในสเปกตรัมการดูดกลืนแสงชั่วคราว (ติดตามการถ่ายโอนอิเล็กตรอนทั้งไปข้างหน้าและข้างหลัง)
• ภายใต้ K ET ≫ K₀(C) การสลายตัวของ ESA ของ C จะสะท้อนถึง K ET (เนื่องจาก ESA หายไปเมื่อ C กลายเป็น C⁺)
• จลนศาสตร์ GSB ของ D สะท้อนทั้ง K ET และ K BET.
• การฟื้นตัวของ GSB ของ C สะท้อนถึง K BET.
2) ทราบสเปกตรัมการดูดกลืนแสงในสถานะคงตัวของ C และอยู่ภายในช่วงการตรวจจับ แต่ไม่ทราบสเปกตรัมของ C⁺ หรืออยู่นอกช่วง ไม่ทราบสเปกตรัมของ D หรือตรวจไม่พบ แต่สเปกตรัมของ D⁻ เป็นที่รู้จักและอยู่ภายในช่วงการตรวจจับ
รูปที่ 6a แสดงสเปกตรัมการดูดกลืนแสงของ C และ D⁻ รูปที่ 6b แสดงสเปกตรัมการดูดกลืนแสงชั่วคราวที่แก้ไขตามเวลา
หลังจากการกระตุ้นของ C สัญญาณการดูดซึม D⁻ จะปรากฏขึ้นและเพิ่มขึ้นตามเวลาหน่วง ซึ่งบ่งชี้ถึงการถ่ายโอนอิเล็กตรอนจาก C ไป D ขณะเดียวกัน GSB ของ C จะไม่สลายตัวตามเวลาภายใต้ K ET ≫ K₀(C) แต่ ESA ของ C จะสลายตัวเนื่องจากการแปลงเป็น C⁺
เนื่องจากเวลาล่าช้ายังคงเพิ่มขึ้น กระบวนการเดิมพันจึงเริ่มมีอิทธิพลเหนือ สัญญาณจาก C และ D⁻ สลายตัว และระบบจะกลับสู่สถานะกราวด์
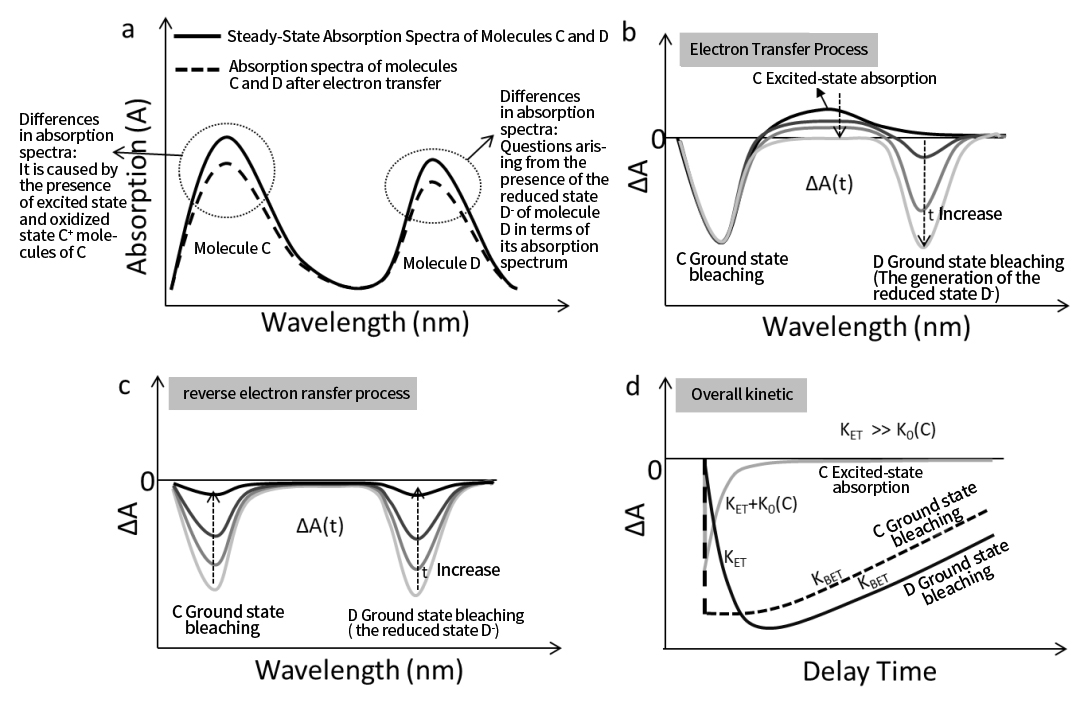
รูปที่ 5 (a) สเปกตรัมการดูดกลืนแสงในสถานะคงตัวของโมเลกุล C และ D ควบคู่ไปกับสเปกตรัมการดูดกลืนแสงชั่วคราวของโมเลกุล C ในช่วงเวลาหนึ่งหลังจากการถ่ายโอนอิเล็กตรอนจาก C ไป D เมื่อถูกกระตุ้น (b) สเปกตรัมการดูดกลืนแสงชั่วคราวที่เวลาหน่วงต่างๆ แสดงให้เห็นกระบวนการถ่ายโอนอิเล็กตรอนด้วยแสงระหว่าง C และ D (c) สเปกตรัมการดูดกลืนแสงชั่วคราวที่เวลาหน่วงต่างๆ แสดงกระบวนการถ่ายโอนอิเล็กตรอนแบบย้อนกลับ (d) เส้นโค้งจลน์ที่ถูกดึงออกมาที่ตำแหน่งต่างๆ ของสเปกตรัมลักษณะการดูดกลืนแสงชั่วคราว
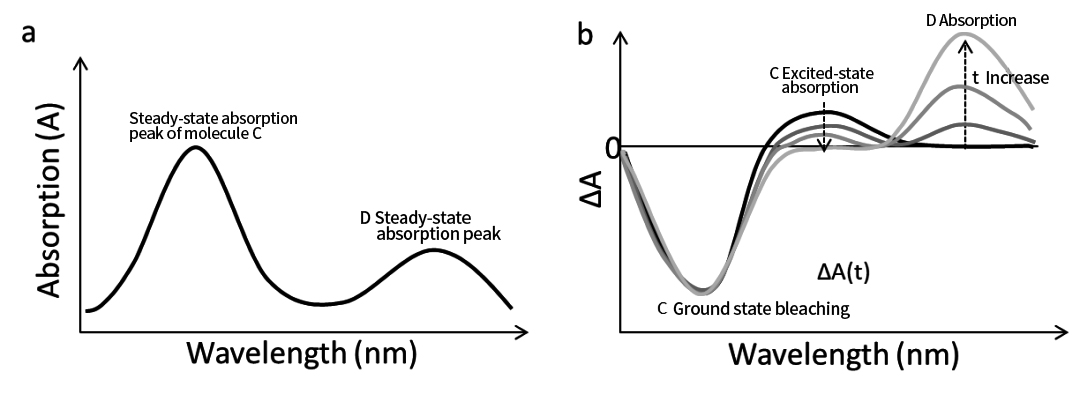
รูปที่ 6 (a) สเปกตรัมการดูดกลืนแสงในสภาวะคงตัวของโมเลกุล C และ D⁻ (b) สเปกตรัมวิวัฒนาการเวลาการดูดกลืนแสงชั่วคราวที่สอดคล้องกันเผยให้เห็นกระบวนการถ่ายโอนอิเล็กตรอนด้วยแสงจากโมเลกุล C ไปยังโมเลกุล D
ในทำนองเดียวกัน หากทราบสเปกตรัมการดูดกลืนแสงของโมเลกุล C⁺ เราก็สามารถสังเกตการเติบโตของสัญญาณการดูดกลืนของมันในสเปกตรัมการดูดกลืนแสงชั่วคราวเมื่อการถ่ายโอนอิเล็กตรอนด้วยแสงเกิดขึ้น
3) ในสถานการณ์ที่สาม ทราบเฉพาะจุดสูงสุดของการดูดกลืนแสงในสภาวะคงตัวของโมเลกุลผู้บริจาคอิเล็กตรอน C เท่านั้น ในขณะที่ไม่ทราบข้อมูลสเปกตรัมของ C⁺, D และ D⁻ และอยู่นอกช่วงการตรวจจับสเปกตรัม
ในกรณีนี้ เราสามารถระบุได้ว่าการถ่ายโอนอิเล็กตรอนเกิดขึ้นหรือไม่โดยการเปรียบเทียบสัญญาณการดูดซึมสถานะตื่นเต้น (ESA) ของโมเลกุล C ภายใต้สภาวะที่มีและไม่มีการถ่ายโอนอิเล็กตรอน
ดังแสดงในรูปที่ 7 ภายใต้เงื่อนไขการถ่ายโอนอิเล็กตรอน การสลายตัวของสัญญาณ ESA ของ C จะเร็วขึ้น นี่เป็นเพราะกระบวนการสลายตัวในสภาวะตื่นเต้นไม่เพียงแต่รวมถึงอัตราการสลายตัวภายใน K₀(C) เท่านั้น แต่ยังรวมไปถึงวิถีการสลายตัวเพิ่มเติมจากการถ่ายโอนอิเล็กตรอน (K ET ) ด้วย
รูปที่ 7b แสดงเส้นโค้งจลน์ของสัญญาณ ESA ของ C ภายใต้ทั้งสองเงื่อนไข เมื่อเปรียบเทียบโปรไฟล์การสลายตัวทั้งสองนี้ จะสามารถคำนวณค่าคงที่อัตราการถ่ายโอนอิเล็กตรอน (KET) ได้
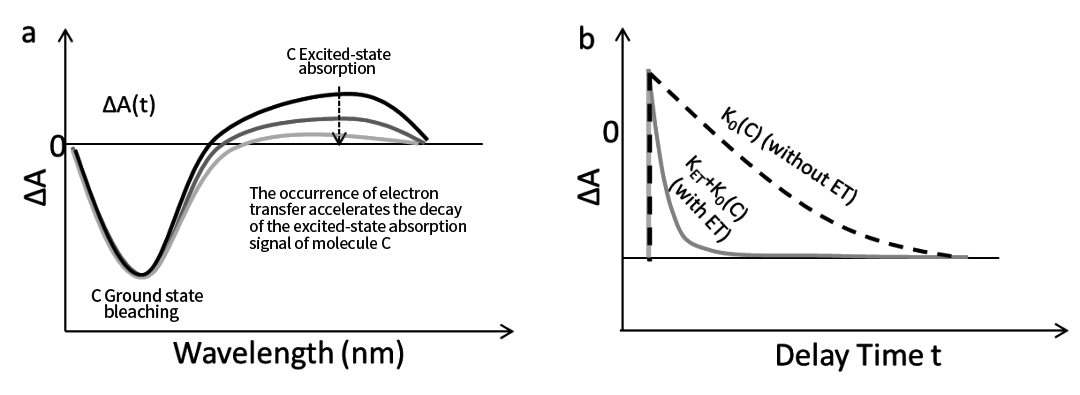
รูปที่ 7 (a) การถ่ายโอนอิเล็กตรอนด้วยแสงช่วยเร่งการสลายตัวของสัญญาณการดูดกลืนสถานะตื่นเต้นของโมเลกุลผู้บริจาค C (b) เส้นโค้งจลนศาสตร์ของสัญญาณการดูดกลืนสถานะตื่นเต้น โดยมีและไม่มีการถ่ายโอนอิเล็กตรอน อัตราจลน์ของการถ่ายโอนอิเล็กตรอนสามารถหาได้โดยการเปรียบเทียบพารามิเตอร์จลน์ของพวกมัน
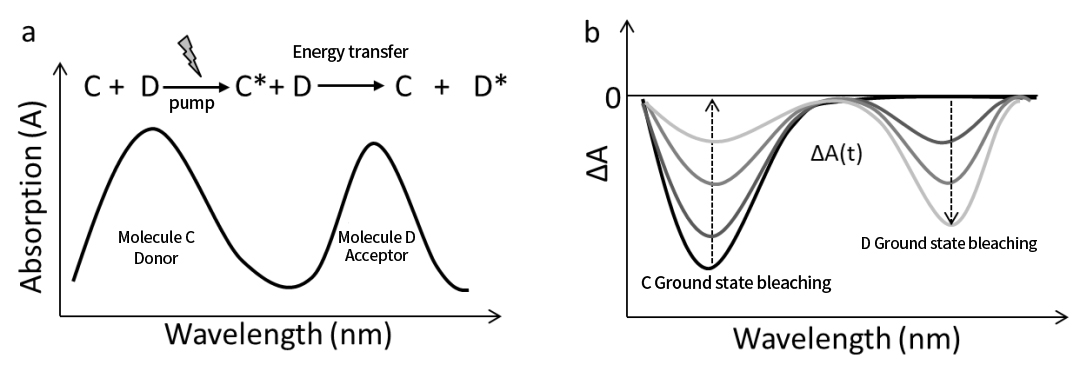
รูปที่ 8 (a) สเปกตรัมการดูดกลืนแสงในสภาวะคงตัวของโมเลกุลผู้ให้พลังงาน C และโมเลกุลตัวรับพลังงาน D ในการถ่ายโอนพลังงานที่เกิดจากแสง ควบคู่ไปกับกระบวนการปฏิกิริยาการถ่ายโอนพลังงาน (b) สเปกตรัมวิวัฒนาการตามเวลาของการดูดกลืนชั่วคราวของโมเลกุล C และ D ในระหว่างกระบวนการถ่ายโอนพลังงาน
04 การถ่ายโอนพลังงานด้วยแสง
นอกจากนี้ สเปกโทรสโกปีการดูดกลืนแสงชั่วคราวยังสามารถใช้เพื่อตรวจจับกระบวนการถ่ายโอนพลังงานที่เกิดจากแสงระหว่างโมเลกุลได้อย่างมีประสิทธิภาพ
ตัวอย่างเช่น รูปที่ 8a แสดงสเปกตรัมการดูดกลืนแสงในสภาวะคงตัวของโมเลกุลผู้ให้ C และโมเลกุลตัวรับ D หลังจากการกระตุ้นด้วยแสง โมเลกุล C จะเข้าสู่สภาวะตื่นเต้น จากนั้นพลังงานกระตุ้นจะถูกถ่ายโอนไปยังโมเลกุล D โดยการถ่ายโอนพลังงาน ทำให้ D รู้สึกตื่นเต้นในสภาวะตื่นเต้น ในขณะที่โมเลกุล C กลับคืนสู่สถานะพื้น
กระบวนการถ่ายโอนพลังงานนี้สะท้อนให้เห็นในสเปกตรัมการดูดกลืนแสงชั่วคราว ดังแสดงในรูปที่ 8b:
• สัญญาณสารฟอกขาวในสถานะพื้นดิน (GSB) ของโมเลกุล C จะสลายตัวหลังจากการกระตุ้น
• ขณะที่สัญญาณ GSB ของโมเลกุล D ค่อยๆ ปรากฏขึ้น แสดงว่า D เริ่มตื่นเต้นแล้ว
การแยกเส้นโค้งจลน์ออกจากสเปกตรัมชั่วคราว ทำให้สามารถกำหนดค่าคงที่ของอัตราการถ่ายโอนพลังงานได้
ดังที่เราเห็น ไม่เหมือนกับการถ่ายโอนอิเล็กตรอนด้วยแสง สัญญาณ GSB ของโมเลกุล C จะสลายตัวในระหว่างกระบวนการถ่ายโอนพลังงาน เนื่องจากสถานะที่ตื่นเต้นของ C จะกลับสู่สถานะพื้นดินอันเป็นผลมาจากการถ่ายโอนพลังงาน
โปรดทราบว่าในรูปที่ 8b สัญญาณการดูดซึมสถานะตื่นเต้น (ESA) ของ C และ D จะไม่แสดง หากสังเกตได้ พฤติกรรมจลนศาสตร์ระหว่างการถ่ายโอนพลังงานจะสะท้อนวิวัฒนาการของสัญญาณ GSB ที่สอดคล้องกัน
05 การปล่อยก๊าซกระตุ้น
นอกจากนี้ สเปกโทรสโกปีการดูดกลืนแสงชั่วคราวยังสามารถใช้เพื่อตรวจจับกระบวนการกระตุ้นการปล่อยก๊าซเรือนกระจก (SE) ของโมเลกุลได้อีกด้วย
กระบวนการ SE เกิดขึ้นจากการเชื่อมโยงกันระหว่างการปล่อยฟลูออเรสเซนซ์ของโมเลกุลและแสงของโพรบภายในช่วงสเปกตรัมเรโซแนนซ์ (ดังแสดงในรูปที่ 10a) โดยเฉพาะอย่างยิ่ง เมื่อแสงจากโพรบมาถึง โมเลกุลบางตัวในสถานะตื่นเต้น S₁ จะมีปฏิกิริยากับโฟตอนของโพรบและปล่อยแสงที่ถูกกระตุ้นออกมา
ตำแหน่งสเปกตรัมของสัญญาณ SE ตรงกับตำแหน่งสเปกตรัมของการเรืองแสงที่เกิดขึ้นเองของตัวอย่าง และโดยทั่วไป SE จะปรากฏบนขอบสีแดงของสัญญาณสารฟอกสีสถานะพื้นดิน (GSB) ในหลายกรณี สัญญาณ SE และ GSB ตั้งอยู่ใกล้กันจนทับซ้อนกันบางส่วน ดังแสดงในรูปที่ 9b
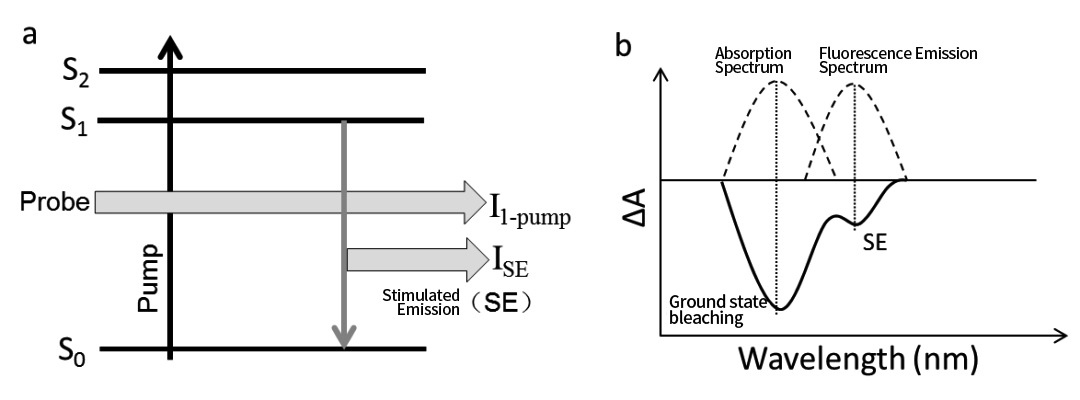
รูปที่ 9 กระบวนการกระตุ้นการปล่อยโมเลกุลและสเปกตรัมการดูดกลืนแสงชั่วคราวที่สอดคล้องกัน
สัญญาณกระตุ้นการปล่อย (SE) ในสเปกโทรสโกปีการดูดกลืนแสงชั่วคราวจะปรากฏเป็นสัญญาณคล้ายสารฟอกขาว (ลบ) เนื่องจากหลังจาก SE เกิดขึ้นที่ความยาวคลื่นการปล่อยแสงฟลูออเรสเซนซ์ ความเข้มของลำแสงโพรบที่ไปถึงเครื่องตรวจจับจะเพิ่มขึ้นเนื่องจากมีการเพิ่มการปล่อยสัญญาณกระตุ้น:
ฉัน 1-ปั๊ม + ISE > ฉัน 1-unpump
เป็นผลให้การเปลี่ยนแปลงการดูดกลืนแสงคำนวณได้ดังนี้:
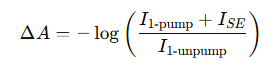
ซึ่งให้ค่า ∆A เป็นลบ ซึ่งคล้ายกับสัญญาณการฟอกขาวในสถานะพื้นดิน (GSB)
อย่างไรก็ตาม สิ่งสำคัญคือต้องสังเกตว่าสัญญาณ SE สะท้อนถึงจำนวนประชากรของโมเลกุลในสถานะตื่นเต้น S₁ ในขณะที่ GSB สะท้อนถึงการพร่องของโมเลกุลสถานะพื้น ตัวอย่างเช่น ในระหว่างกระบวนการถ่ายโอนอิเล็กตรอนในสถานะ S₁ สัญญาณ SE จะสลายตัวอย่างรวดเร็ว (เมื่อประชากร S₁ ลดลง) ในขณะที่สัญญาณ GSB ยังคงไม่เปลี่ยนแปลง (เนื่องจากโมเลกุลในสถานะพื้นยังไม่ฟื้นตัว)
06 บทสรุป
บทสรุปของกระบวนการจลน์ศาสตร์ในสภาวะตื่นเต้นที่สำคัญในระบบโมเลกุลและสัญญาณสเปกตรัมชั่วคราวที่สอดคล้องกันของกระบวนการดังกล่าวได้แสดงไว้ในรูปที่ 10 ดังที่แสดงให้เห็นแล้ว สเปกโทรสโกปีการดูดกลืนแสงชั่วคราวสามารถตรวจจับกระบวนการในสถานะตื่นเต้นที่สำคัญที่สุดในระบบโมเลกุลได้ อย่างไรก็ตาม สัญญาณจลน์ศาสตร์ต่างๆ เหล่านี้มักจะทับซ้อนกันและเกี่ยวพันกัน ซึ่งทำให้เกิดความท้าทายในการตีความทั้งสเปกตรัมชั่วคราวและข้อมูลจลน์ศาสตร์
ในการทดลองจริง จำเป็นต้องมีการวิเคราะห์อย่างรอบคอบ สามารถใช้เทคนิคต่างๆ เช่น การปรับให้เหมาะสมโดยรวม การสร้างแบบจำลองจลน์ศาสตร์ และการทดลองควบคุมเพื่อกำหนดและยืนยันต้นกำเนิดของสัญญาณชั่วคราวต่างๆ
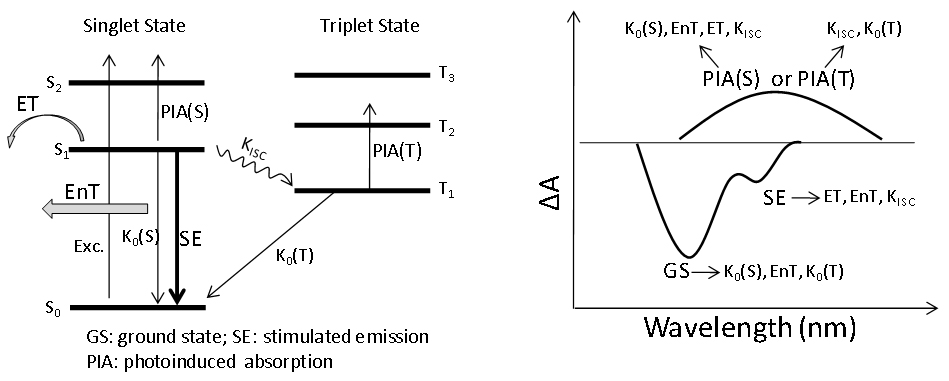
รูปที่ 10 กระบวนการไดนามิกในสภาวะตื่นเต้นหลักและคุณลักษณะทางสเปกตรัมที่สอดคล้องกันซึ่งสามารถตรวจพบได้โดยสเปกโทรสโกปีการดูดกลืนแสงชั่วคราวของระบบโมเลกุล ลูกศรในรูปด้านขวาแสดงถึงกระบวนการไดนามิกที่เป็นไปได้ของสัญญาณสเปกตรัม
(สงวนลิขสิทธิ์ กรุณาอ้างอิงแหล่งที่มา)